

ミニ特集「身体の情報処理がこころをつくる」:社会的認知発達における身体の役割を探る:身体の外側と内側の視点から
心理・認知科学専攻 平井真洋
私たちは日常生活で,日々多くのヒトに囲まれ,多くのヒトの動きを目にしています.このような他者の動きを理解することが,社会生活には不可欠であることに異論はないでしょう.みなさんは友人・知人の何気ない仕草から相手の考えを読み解こうとした経験が数多くあると思います.そういった私たちがいかに他人の動き(所作)に敏感であるかの例をまずはご覧ください↓.
わずか十数個の光点運動が表示されているだけにもかかわらず,ありありとヒトが動いている様を感じることができることに気づかれたと思います.これはモーションキャプチャー装置と呼ばれる動きを精密に計測可能な装置によって作られ,各光点はヒトの関節や頭部といった主要な箇所にのみ装着されたマーカーの位置から生成されています.ています.みなさんもテレビゲームや映画で実在するヒトの様に動くCGキャラクターを目にすることが多いと思います.そのうちいくつかはヒトが演じた動きをモーションキャプチャー装置により生成されているはずです.最近ではそのような大型装置を使わなくとも,Openposeと呼ばれる自然動画からヒトの動きを自動的に切り出すことができる深層学習を使ったプログラムにより,お手軽にヒトの動きを生成できる時代になっています.10年前には想像できない世界が今,広がっているのです.
話を戻します.このような知覚現象は「バイオロジカルモーション」と呼ばれ,1970年代にスウェーデンの心理学者グンナー・ヨハンソンによって報告されました(Johansson, 1973).その後現在に至るまで約50年にも渡り心理学者を魅了し続け,この十数個の光点運動のみから実に様々な情報を読み取ることが可能であることが解明されつつあります.
これまでの多くの研究成果の蓄積により,バイオロジカルモーションから進行方向やヒトの形といった情報だけでなく,性別,意図,感情,自分か他人か,騙そうとする動きかどうか,怪しい動きかどうか,重たい荷物を持つように演技しているかどうか(軽い荷物を持ち上げているにもかかわらず,重たい荷物を持ち上げるような演技を見破れるか)など他者に関する情報を読み取ることが可能であることがわかっています.
よくよくこの知覚現象を考えると本当に不思議です.ヒトの外見に関する情報を取り除き,ヒトの動きに関する情報についても極限まで削ぎ落とした動きにもかかわらず,私たちの視覚系はヒトに関する極めて豊かな情報を「再構築」することができるからです.
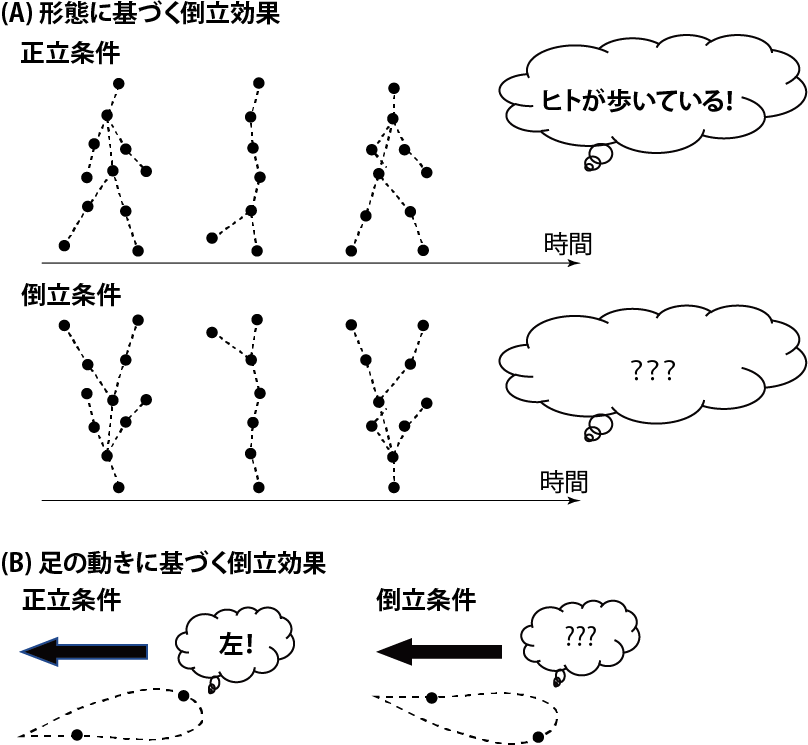
この現象をどのように理解すれば良いのでしょう?一見するととても複雑な現象のように見えますが,ヒトが歩いている際のバイオロジカルモーションに限定すると,この現象は二つの「倒立効果」から理解できそうです.
一つ目の倒立効果(図1A)とは,顔の知覚に見られる倒立効果(顔を180度ひっくり返すと顔がわかりにくくなる効果です)と類似の効果です(Sumi, 1984).つまり,歩行のバイオロジカルモーションを180度ひっくり返すともはやそこには歩いているヒトは見当たらず,例えば単なる竜巻のような点の動きの集まりにしか見えない,と報告する方もいます.二つ目の倒立効果(図1B)とは,ヒトの形とは独立の「足の動き」に基づく倒立効果です(Troje & Westhoff, 2006).これはバイオロジカルモーションの各光点を空間的にランダムに配置し,足の運動軌跡を180度ひっくり返すことで,どちらに進んでいるのかがわからなくなってしまう現象です.足の動きが正立であれば,バイオロジカルモーションの各光点の空間的位置をランダマイズした場合であっても(そこにはもはやヒトの形は見えないのですが),左右どちらに歩いているかを正確に判断することができます.これは,足の動きそれ自体に進行方向に関する情報が埋め込まれていることを示しています.足の動きは筋骨格系の制約だけでなく,重力によっても影響を受けています.ヒトが歩く際にはこの二つの制約によって生成さえる特徴的な動きがあり,私たちの視覚系はそのような特徴的な情報を足の動きから解読している可能性があります.このようにバイオロジカルモーションには点の動きから知覚される形態情報に関する処理と,足の動きのような局所的な動きの情報処理が関与する現象であると言えそうです.
私たちがこれほどまでにヒトの動きに敏感であるということは,脳の中には「ヒトの動き」の検出に特化した神経回路があると思われた方もおられるのではないでしょうか.実際,まさにそのとおりで,私たちの脳の中にはバイオロジカルモーション,もう少し広く言うならば,他者の動作(行為)や他者の情報関して選択的に活動する場所があることが知られています.
専門的な話になりますが,バイオロジカルモーションを観察している際には,上側頭溝(Superior Temporal Sulcus)と呼ばれる,耳の少し上のあたりに位置する脳が選択的に活動することが知られています(Grossman & Blake, 2002).さらに,バイオロジカルモーションの処理は脳内において,映像を見てから300〜500ミリ秒内という短時間で完了している可能性を私たちの研究グループは見出しつつあります(Hirai, Fukushima, & Hiraki, 2003; Hirai & Kakigi, 2008; Hirai, Kaneoke, Nakata, & Kakigi, 2008).興味深いのは,バイオロジカルモーションの観察には視覚的な処理だけではなく,運動を司る場所も関与する可能性が報告されているということです(Saygin, 2007; Saygin, Wilson, Hagler, Bates, & Sereno, 2004).これは後でも触れますが,他人の動きを見ることは見る処理と同時に自分自身の動きを制御する脳の場所も同時に活動することを意味します.実はこの上側頭溝領域はとても興味深い場所で,ほかにも他人の視線や表情などを観察する場合でも活動することが知られています.つまり,他人に関する情報をこの場所で処理しているとも言えそうです.実際,私たちの脳は「社会的コミュニケーションを取るために設計された」のではないかとする「社会脳仮説」があります(Brothers, 1990).
社会脳仮説のメガネを通して脳の働きをみると,脳の至るところに他者に関する情報処理に選択的に反応する場所あります.例えば,顔を見た際に選択的に活動する「紡錘状回顔領域(Fusiform Face Area)」や他者の心の状態を推測する際に選択的に活動する「側頭頭頂接合部(Temporoparietal junction)」,ヒトの身体の静止画に選択的に活動する「外線条体身体領域(Extrastriate Body Area)」など,これ以外にも複数存在します(脳部位の具体的な場所は図2をご覧ください).
実際の脳の場所はこちらをご参照下さい:
- 紡錘状回顔領域: https://ja.wikipedia.org/wiki/%E7%B4%A1%E9%8C%98%E7%8A%B6%E5%9B%9E
- 側頭頭頂接合部: https://ja.wikipedia.org/wiki/%E5%81%B4%E9%A0%AD%E9%A0%AD%E9%A0%82%E6%8E%A5%E5%90%88%E9%83%A8

(話は脱線しますが,収束の気配がまだ見えない,新型コロナウィルス感染症ももしかしたら「社会脳」が持っている「他者とコミュニケーションしたい根源的な欲求」のチャネルにとりついた狡猾なウィルスなのかもしれません.)
このような脳の活動場所を調べた研究は,脳の機能がある程度完成された成人を対象とした研究結果に基づくものです.また,勘の鋭い読者の方ならこう思うかもしれません.「いつから私たちはバイオロジカルモーションのような他者の動きに関する情報処理ができるのか」と.
実際,バイオロジカルモーションの発達研究は1980年代より始まり,驚くべきことに,視覚経験がほぼなく,視力がまだ十分発達していない,生まれて十数時間の赤ちゃんでも,バイオロジカルモーションとランダムに動く光点を区別できることが今から13年ほど前に報告されています(Simion, Regolin, & Bulf, 2008).実はバイオロジカルモーションだけに限らず,生まれてすぐの赤ちゃんでも顔とそうでないもの,アイコンタクトがある顔とそうでない顔を区別することができることが知られています.これは,何を意味するのでしょうか?一つの可能性として考えられることは,ヒトの動きや顔といったいわゆる「社会的な刺激」を検出する仕組みが生まれながらに備わっている可能性がある,ということです.
バイオロジカルモーションに限定して考えると,バイオロジカルモーションの知覚発達のシナリオとして,上述の二つ目の倒立効果で示したようなヒトの足の動きが重要な役割を果たすと私たちの研究グループは考えています.つまり,足の動きが新生児の注意を惹きつけ,これにより新生児はヒトの動きに関する情報入力が増加し,大脳皮質上にバイオロジカルモーション(他者の動き)に選択的な領域が形成される,と想定しています.これは,1990年代に提唱された顔認知の発達過程に関する理論モデル(Morton & Johnson, 1991)をお手本としてヒトの動きの知覚発達をモデル化したものです(Hirai & Senju, 2020).もちろんこのモデルはバイオロジカルモーション知覚発達の一つのシナリオに過ぎませんが,様々な研究成果を勘案すると,このシナリオのような発達過程をたどる可能性は高いのではないかと考えています.
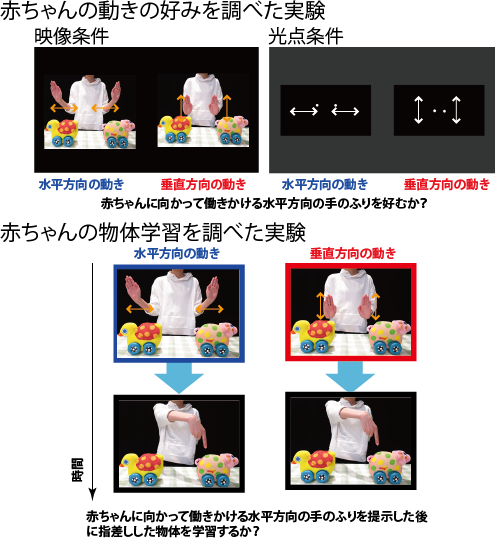
また,乳児は他者の動きを検出するだけでなく,他者の動きの情報を利用して学びにつなげることがわかりつつあります.例えば養育者が乳児に向かって何かを教えようとする際,養育者は乳児に対して大げさな繰り返しの動き(モーショニーズ)を示すことが報告されています(Brand, Baldwin, & Ashburn, 2002).確かに,モーショニーズは乳児の注意をひきつけますが(Brand & Shallcross, 2008),大きな手の動き,繰り返しの動きは物理的な顕著性が高いため,乳児の注意をひきつけるのは当然かもしれません.私たちは,動きに潜む乳児の学びを促す手がかりについても研究を始めています(図3).詳細は省きますが,赤ちゃんに向かって働きかける物理的特徴量としては同じ二つの動き,すなわち,横にふった手の動きと縦にふった手の動きを同時に提示した場合,横に振った手の動き(みなさん誰かに働きかけるときにするあの手の動きです)は縦に振った手の動きよりも乳児の注意をひきつけ,手をふったあとに指差しする玩具をよく記憶できることがわかりました(Hirai & Kanakogi, 2019).他にも,どのような身体の動きが乳児の注意をひきつけ,物体の学習を促進するかに関する研究を現在進めています(Hirai et al., in press).
このように私たちは生まれながらに他人の動きに敏感になるように設計されているのです.そして,他者に囲まれて育つことによって私たちはヒトの動きを読み解くプロフェッショナルとなり,その手がかりをうまく使うことによって世界に関する知識を増やしていく可能性が考えられます.
身体の内側:自己に敏感な私たちの脳―「人の立場にたって」を科学する
前節では,私たちの脳が他者に敏感となるように設計されていること,そしてヒトの動き(バイオロジカルモーション)を観察したときには,視覚的な処理だけではなく,運動制御に関連する脳部位も活動することを紹介しました.これは,他人の動きがあたかも自分の動きのように「ミラー」され,脳の中で処理されている可能性を示しています(Rizzolatti & Craighero, 2004).このように脳の中には自己に関する情報処理,他者に関する情報処理そしてその両方の情報処理を担う場所があり,この自己の動きと他者の動きの処理が重なり合う脳の部位は「ミラーニューロンシステム」と呼ばれています.
自己・他者の情報処理はもちろん他人の動き・自分の動きだけに限ったことではなく,相手がどのようにものを見ているのか,という場面でも当てはまります.みなさんもおそらくは小さい頃,「他の人の立場に立って物事を進めなさい」「他の人の気持ちになって」と親御さんから言われたことがあるのではないでしょうか?これはまさに相手の心の状態をシミュレーションすることを親御さんから求められているわけです.では,どうやって私たちは相手の立場に立って物事(相手の視点)を考える能力を身につけるのでしょうか?
「幽体離脱体験」という言葉をお聞きになったことのある読者の方は多いと思います.決してオカルトの話ではなく,科学的にこれまで研究がなされています.例えば検査のために脳内に電極を埋め込んだ患者さんの脳の頭頂側頭接合部(Temporoparietal junction)を刺激すると,自分自身が自分の身体から離れてしまう感覚に陥るとOlaf Blankeらのグループは報告しています(Blanke & Arzy, 2005).つまり,自分の身体感覚が脳の特定の領域を刺激操作することによって容易に変容するわけです.おそらくみなさんは自己身体の感覚はとても頑健でゆるがないと思われていると思いますが,いとも簡単に自己身体の感覚は変容してしまいます(ラバーバンドイリュージョンと呼ばれる現象(Botvinick & Cohen, 1998)もまさにそのような現象を端的に,実験的に示したものです https://www.youtube.com/watch?v=sxwn1w7MJvk).
このような頭頂側頭接合部は,他人はどう見えているのか,という相手の視点を考える場合にも活動することが知られています.これはつまり,「自分自身の身体」を他人のいる場所に空想して移動する操作が脳内において行われている可能性を示しています.つまり,自分自身の身体を「操作」して,相手の位置に自分自身を移動することをシミュレーションすることで,相手の視点を理解しようとする可能性があります.私たちは,心的回転課題と他者視点取得課題をツールとして用いることによりこの問題に取り組んでいます(図4).このような能力が成人に近づくのがおよそ11〜12歳ぐらいではないか,というのが私たちの研究グループの見立てです(Hirai, Muramatsu, & Nakamura, 2020).

図4.(A)心的回転課題と(B)他者視点取得課題(上から見た図).(C)4歳から12歳までの児を対象とした二つの課題の課題成績(Hirai et al., 2020).
このように私たちは他人の動きから様々な情報を読み取り,相手の状態を推定し,かつ,自分の身体を巧みに利用することにより,相手の状態をシミュレーションすることが可能なようです.しかしながら,それらがどのように発達変化するか,どのような要因がいつどのように影響するのかについては未だ多くの謎に包まれています.また,定型発達の方だけではなく,非定型発達の方々も特徴的な発達過程を示すことが多くの報告により明らかにされています.私たちの研究グループでは,赤ちゃんから成人まで,様々な発達過程を持っておられる方を対象にこのような身体に根ざした社会的認知発達の研究を進めており,これからも様々な方法論を有機的に組み合わせてそのメカニズムを明らかにしていきます.ご興味のある方は以下のリンクからホームページにご訪問下さい(平井研究室ホームページ: https://www.hirailab.cog.i.nagoya-u.ac.jp/index.html)
参考文献
Blanke, O., & Arzy, S. (2005). The out-of-body experience: disturbed self-processing at the temporo-parietal junction. Neuroscientist, 11(1), 16-24.
Botvinick, M., & Cohen, J. (1998). Rubber hands ‘feel’ touch that eyes see. Nature, 391, 756.
Brand, R. J., Baldwin, D. A., & Ashburn, A. L. (2002). Evidence for ‘motionese’: modifications in mothers’ infant-directed action. Developmental Science, 5(1), 72-83.
Brand, R. J., & Shallcross, W. L. (2008). Infants prefer motionese to adult-directed action. Dev Sci, 11(6), 853-861. doi:10.1111/j.1467-7687.2008.00734.x
Brothers, L. (1990). The social brain: a project for integrating primate behavior and neuropsychology in a new domain. Concepts in Neuroscience, 1, 27-51.
Grossman, E. D., & Blake, R. (2002). Brain Areas Active during Visual Perception of Biological Motion. Neuron, 35(6), 1167-1175.
Hirai, M., Fukushima, H., & Hiraki, K. (2003). An event-related potentials study of biological motion perception in humans. Neuroscience Letters, 344(1), 41-44.
Hirai, M., & Kakigi, R. (2008). Differential cortical processing of local and global motion information in biological motion: an event-related potential study. J Vis, 8(16), 2 1-17.
Hirai, M., & Kanakogi, Y. (2019). Communicative hand-waving gestures facilitate object learning in preverbal infants. Dev Sci, 22(4), e12787. doi:10.1111/desc.12787
Hirai, M., Kaneoke, Y., Nakata, H., & Kakigi, R. (2008). Neural responses related to point-light walker perception: a magnetoencephalographic study. Clinical Neurophysiology, 119(12), 2775-2784.
Hirai, M., Muramatsu, Y., & Nakamura, M. (2020). Role of the Embodied Cognition Process in Perspective-Taking Ability During Childhood. Child Development, 91(1), 214-235. doi:10.1111/cdev.13172
Hirai, M., & Senju, A. (2020). The two-process theory of biological motion processing. Neuroscience and Biobehavioral Reviews, 111, 114-124. doi:10.1016/j.neubiorev.2020.01.010
Hirai, M., Kanakogi, Y., & Ikeda, A., (in press).
Johansson, G. (1973). Visual perception of biological motion and a model for its analysis. Perception and Psychophysics, 14, 201-211.
Morton, J., & Johnson, M. H. (1991). CONSPEC and CONLERN: a two-process theory of infant face recognition. Psychological Review, 98(2), 164-181.
Rizzolatti, G., & Craighero, L. (2004). The mirror-neuron system. Annual Review of Neuroscience, 27, 169-192.
Saygin, A. P. (2007). Superior temporal and premotor brain areas necessary for biological motion perception. Brain, 130(Pt 9), 2452-2461.
Saygin, A. P., Wilson, S. M., Hagler, D. J., Jr., Bates, E., & Sereno, M. I. (2004). Point-light biological motion perception activates human premotor cortex. Journal of Neuroscience, 24(27), 6181-6188.
Simion, F., Regolin, L., & Bulf, H. (2008). A predisposition for biological motion in the newborn baby. Proceedings of the National Academy of Sciences of the United States of America, 105(2), 809-813.
Sumi, S. (1984). Upside-down presentation of the Johansson moving light-spot pattern. Perception, 13(3), 283-286.
Troje, N. F., & Westhoff, C. (2006). The inversion effect in biological motion perception: evidence for a “life detector”? Current Biology, 16(8), 821-824.